Det vore slöseri med energi för cellen, om alla gener ständigt uttrycktes, och det skapades en mängd proteiner, som inte behövs för tillfället.
Därför behövs det en reglering av transkriptionen!
Hushållsgener uttrycks konstitutivt
Vissa gener (ett fåtal) är inte reglerade på annat sätt än att de har olika effektiva promotorer
De uttrycks konstitutivt (hela tiden).
Kallas ibland hushållsgener, eftersom de behövs hela tiden för att sköta grundläggande funktioner i cellen.
Andra gener (de flesta) uttrycks bara när de faktiskt behövs.
Regleringen sker framför allt i initieringen av transkriptionen
Har en gen redan börjat transkriberas, blir det med mycket stor sannolikhet ett färdigt protein
Kan man stoppa transkriptionen innan den börjar, blir regleringen effektivast.
Tre faktorer påverkar initieringen av transkription:
1. Specificitetsfaktorer
Dessa faktorer reglerar RNA-polymerasets specificitet för en viss promotor, d.v.s. hur pass väl RNA-polymraset kan binda till promotorn.
Så är t.ex. fallet med hushållsgenernas promotorer.
2. Repressorer
En repressor är ett protein, som binder till en specifik sekvens i promotorn.
- Denna specifika sekvens kallar man för operatorn.
Genom att fysiskt sitta i vägen, förhindras RNA-polymeraset att börja transkribera genen.
Repressorer utövar negativ reglering – de fungerar som "bromsar".
3. Aktivatorer
En aktivator binder intill eller nära promotorn, och underlättar för RNA-polymeraset att börja transkriptionen.
Ofta är promotorn utformad så, att RNA-polymeraset binder mycket dåligt till den utan hjälp av en aktivator.
- Utan aktivator transkriberas sålunda inte genen, eller i alla fall mycket lite.
Aktivatorer utövar positiv reglering – de "gasar".
Operon
Många prokaryota gener är arrangerade i s.k. operon (se nedan).

- Enzymer består ofta av flera subenheter - flera proteiner - som samverkar. I många fall samverkar också olika proteiner eller enzymer i en kedja för att nå ett gemensamt mål
- Om inte alla subenheterna är med, fungerar inte enzymet. Eller så kan det vara så att om inte alla enzymerna är med i en reaktionskedja, kan inte slutprodukten framställas.
- Det vore oekonomiskt för cellen om t.ex. tre av fyra subenheter transkriberades, men inte den fjärde!
- Ett "smart" sätt som detta lösts på, är att gener som samverkar, ofta sitter omedelbart efter varandra i genomet, och delar på en och samma promotor.
Ett operon kan definieras som en sträcka DNA med en eller flera besläktade gener samt de operator- och promotorsekvenser som reglerar deras transkription.
Lac-operonet
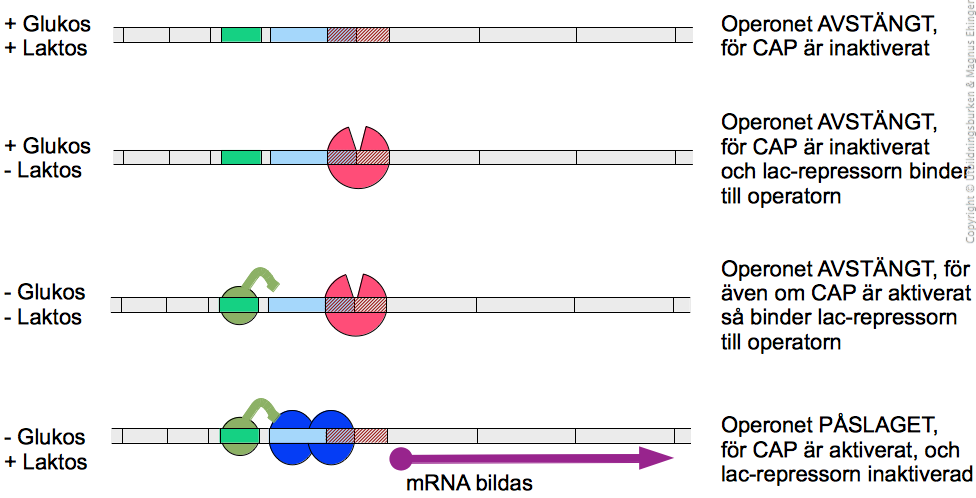
Lac-operonet i E. coli är ett exempel på ett operon, som regleras både positivt och negativt.
På grundläggande nivå, tas lac-operonet ofta upp som ett exempel på reglering av transkription. Tyvärr tar man då bara upp den negativa regleringen, vilket, som vi skall se, bara är halva sanningen.

I lac-operonet (ovan) ingår tre gener som hjälper bakterien att leva på laktos (mjölksocker) som enda kol- och energikälla (se nedan):
- Genen för β-galaktosidas, som spjälkar laktos (LacZ).
- Genen för galaktosid-permeas (LacY). Detta protein bildar en por i cellmembranet, och släpper in laktosmolekylen genom den poren.
- Genen för tiogalaktosid-transacetylas (LacA). Detta proteinets fysiologiska funktion är okänd.
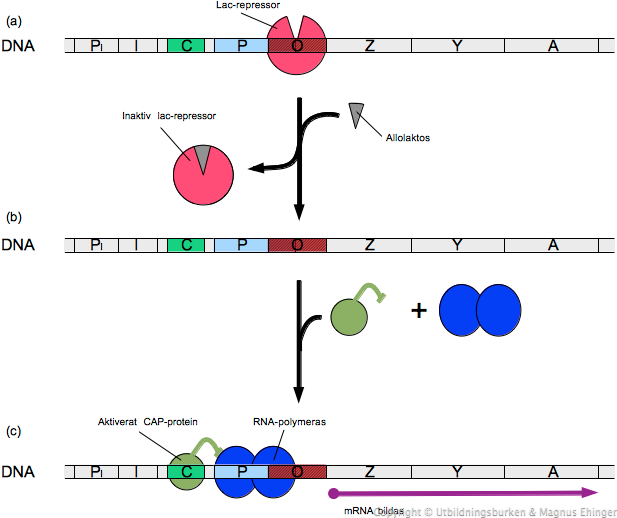
Promotorn (P) och operatorn (O) ingår också i operonet, liksom aktivatorns bindningsställe (C). Slutligen ingår också en repressorgen (I) och dess promotor (PI) i operonet.
Negativ reglering av lac-operonet
När en E. coli-kultur odlas på rikt medium eller glukosinnehållande medium, behöver den inte bryta ner laktos. Då ska lac-operonet alltså vara avstängt.
I-genen uttrycks konstitutivt, och när mRNA:t från denna gen translateras, bildas ett protein som kallas Lac-repressorn. Detta protein finns alltså alltid närvarande.
Lac-repressorn binder in till O-platsen (operatorn) på lac-operonet, och hindrar därmed transkription av lacZ, -Y och -A (se nedan, figur a). Operonet är avstängt.
Allolaktos är en variant av laktos som finns i mycket låga koncentrationer om cellen får växa på laktosinnehållande medium. När allolaktosen binder till lac-repressorn, förändras dess struktur. Repressorn inaktiveras, och släpper från DNA:t. Nu kan DNA:t börja transkriberas till mRNA (se nedan, figur b). Man säger att i detta fallet fungerar allolaktos som induktor, eftersom det gör att repressorn ändrar form (och inaktiveras).
Men här finns en hake: Om bakterien får växa på ett medium som innehåller både glukos och laktos, vad gör den då? Glukos är definitivt den kolkälla bakterien föredrar, så det är onödigt (och slöseri med energi) för bakterien att försöka bryta ner laktos, nu när det finns glukos i överskott. Det är dags för lite positiv reglering!
Positiv reglering av lac-operonet
I cellen finns ett protein, CAP (eng. Catabolite Activator of Proteins), som aktiveras när glukoshalten i cellen börjar sjunka.
Det aktiva CAP-proteinet binder in till C-platsen i lac-operonet (se ovan, figur c). Nu underlättar den för RNA-polymeraset att binda till promotorn, DNA:t kan börja transkriberas, och mRNA kan bildas.
Det är alltså endast när laktoshalten är hög samtidigt som glukoshalten är låg, som lac-operonet är påslaget, och generna lacZ, -Y och -A börjar transkriberas (se nedan). På så vis hushåller E. coli med sina resurser, och tillverkar inte några protein i onödan.