DNA-replikationen följer tre regler
DNA-replikationen följer tre regler
När en cell ska dela sig och bli två, måste arvsmassan kopieras. Detta kallas för att DNA:t i cellen replikeras.
DNA-replikationen följer tre regler:
- Den är semi-konservativ
- Den börjar alltid på samma ställen
- Den sker alltid i 5'→3'-riktning
1. DNA-replikering är semikonservativ.
Den ena DNA-strängen tjänar som mall för syntes av den andra strängen
I replikationen bevaras den ena strängen, och en ny tillverkas.
Eftersom den ena strängen alltid bevaras, är replikationen semi-konservativ = "till hälften bevarande"
Detta föreslogs först av Watson & Crick, och bevisades vara sant av Matthew Meselson och Franklin Stahl i ett mycket elegant experiment år 1957.
- (Fråga eleverna: När var det nu Watson & Crick löste DNA:ts struktur?)
- 1953
Meselsons & Stahls experiment
- E. coli fick växa i många generationer i ett medium, vars enda kvävekälla var NH4Cl, innehållande isotopen 15N.
- DNA isolerades från bakterierna, och visade sig ha c:a 1% högre densitet än "vanligt" DNA (figur (a), nedan).
- Några E. coli överfördes till medium som endast innehöll 14N, och tilläts fördubbla sig endast en ytterligare gång.
- DNA som nu isolerats hade högre densitet än "vanligt" DNA, men lägre än [15N]DNA:t.
- Uppenbarligen hade här bildats [14N, 15N]hybrid-DNA.
- Detta var det egentliga beviset på att replikationen av DNA är semikonservativ (figur (b), nedan).
- Några E. coli överfördes till färskt medium, som endast innehöll 14N, och tilläts fördubbla sig ytterligare en gång.
- DNA som nu isolerats och centrifugerats i en CsCl-gradient, skilde sig i två band: Ett som endast innehöll [14N]DNA och ett som innehöll [14N, 15N]hybrid-DNA (figur (c), nedan).

2. DNA-replikationen börjar alltid på samma ställen
Det ställe där DNA-replikationen börjar kallas origin (ORI).
Från startpunkten sprider sig replikationen i båda riktningarna (bidirektionellt)
En ursprungspunkt (origin) är inte bara var som helst i kromosomen, utan alltid en och samma punkt.
Detta upptäcktes av Ross Inman och hans medarbetare.
- DNA från fag λ denaturerades partiellt.
- Vissa A=T-rika regioner denaturerades alltid först.
- Dessa fick fungera som riktmärken i DNA-molekylen
- Vid replikering kunde Inman se partiellt denaturerat DNA där de inte förväntade sig.
- Det replikerande DNA:t hade alltid samma plats i förhållande till de övriga riktmärkena i molekylen.
Sålunda: Replikering startar alltid på ett bestämt (eller flera bestämda) ställe(n) i genomet.
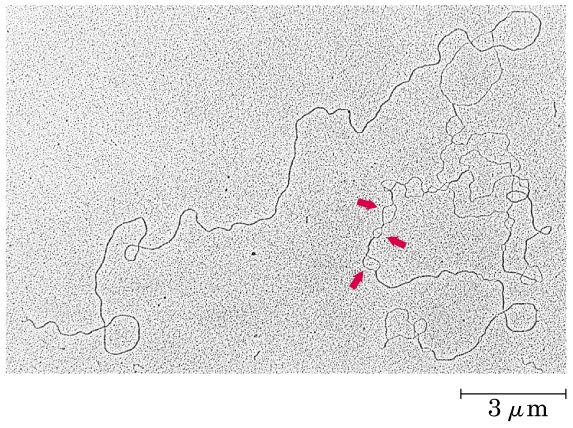
Undersökningar av replikation av cirkulärt DNA visar att replikationen sker i två riktningar (bidirektionellt) (figur 3).

3. DNA-syntes sker i 5'→3'-riktning och är semi-avbruten.
Att DNA-syntes sker i 5'→3'-riktning betyder att på den nysyntetiserade strängens 3'-kol sätts en ny nukleotidgrupp på.
Eftersom DNA är antiparallellt läses mallsträngen av i 3'→5'-riktning.
Detta innebär också med nödvändighet att replikeringen är bidirektionell.
En animering av DNA-replikationen (öppna i nytt fönster):
Om nu DNA-syntes endast sker i 5'→3'-riktning, hur kan då båda strängarna kopieras samtidigt?
Eftersom DNA är antiparallellt, borde en sträng syntetiseras i 5'→3'-riktning och den andra i 3'→5'-riktning. Men det gör den inte!
Detta problem löstes av Reiji Okazaki och hans medarbetare på 1960-talet.
Den ena strängen, den s.k. ledande strängen (eng. leading strand) syntetiseras i ett svep, utan avbrott.
På den andra strängen, den s.k. släpande strängen (eng. lagging strand), syntetiseras nytt DNA i korta stycken, som i efterhand binds samman.
- Dessa korta stycken kallas idag ofta för Okazaki-fragment.
DNA-syntesen är alltså sammanhängande på ena DNA-strängen och avbruten på den andra. Man säger att DNA-syntesen är semi-avbruten.
Okazaki-fragment varierar i storlek från några hundra till några tusen nukleotider, beroende på celltyp.